{kind=link}
Y-STR单倍型在大家系中的差异研究
[张广峰 , 高珊, 畅晶晶, 徐小玉, 郝金萍, 杨雪莹, 朱典, 张颖, 张瑾, 凃政, 刘开会
, 高珊, 畅晶晶, 徐小玉, 郝金萍, 杨雪莹, 朱典, 张颖, 张瑾, 凃政, 刘开会* ]
, 高珊, 畅晶晶, 徐小玉, 郝金萍, 杨雪莹, 朱典, 张颖, 张瑾, 凃政, 刘开会]
|
|


第一作者简介:张广峰(1987—),男,山东泰安人,硕士,主检法医师,研究方向为法医遗传学。E-mail:zhangguangfenga@163.com
, GAO Shan, CHANG Jingjing, XU Xiaoyu, HAO Jinping, YANG Xueying, ZHU Dian, ZHANG Ying, ZHANG Jin, TU Zheng, LIU Kaihui
近年来, Y-STR检验在法医DNA领域的应用日益广泛, 特别是利用Y-STR单倍型进行家系排查在侦查破案中发挥了重要作用。如果不考虑突变因素, Y-STR单倍型在遗传过程中将会保持不变, 这便是家系排查工作的理论基础。然而, Y-STR基因座的突变现象常会给父系亲缘关系的鉴定和家系排查工作带来困扰, 特别是在当前检验的Y-STR基因座数量日益增多, 并且包含较多的高突变率Y-STR基因座的情况下尤甚。研究表明, 不同的Y-STR基因座具有不同的突变率, 常用的Y-STR基因座的平均突变率在2‰ ~3‰ 左右[1]。Ballantyne K等[2]发现DYF387S1、DYS399S1、DYF403S1、DYF404S1、DYS449、DYS518、DYS526b、DYS547、DYS570、DYS576、DYS612、DYS626和DYS627等13个Y-STR的突变率在10‰ 以上, 故称其为快速突变 Y-STR(Rapidly Mutating, RM Y-STR)基因座。目前, 对于Y-STR突变率的调查和Y-STR单倍型在不同家系中的多样性研究较多[2, 3, 4, 5], 而对于同一家系中不同男性个体之间的Y-STR单倍型变化程度研究则较少[6, 7]。可是, 父系亲缘关系的鉴定和家系排查工作都需要对家系中Y-STR单倍型的变化程度有整体上的认知。
本研究选取一遗传关系清晰的汉族家系, 利用Yfiler® Plus复合扩增试剂盒(包含16个YfilerY-STR基因座和DYF387S1、DYS449、DYS518、DYS570、DYS576、DYS627等6个RM Y-STR基因座)和另一包含DYS399S1、DYF403S1、DYF404S1、DYS526b、DYS547、DYS612、DYS626等7个RM Y-STR基因座的复合扩增体系, 获得了所采样本的32个Y-STR基因座的分型数据, 并比较了Yfiler、YfilerPlus和RM Y-STR单倍型在世代遗传过程中的变异程度的差异, 希冀为父系亲缘关系鉴定和家系排查提供参考。
选取一遗传关系清晰的汉族家系(图1), 按照知情同意的原则, 共采集到其中47名男性个体的口腔拭子样本。样本编号以代数+序号表示, 如8-1表示家系中的第八代第一名个体。由于家系图中部分个体的样本未能采集, 故样本编号存在有不连续的现象。家系中两两个体之间的亲缘距离为1次减数分裂(父子对, 此处指产生后代的实际显效减数分裂)到19次减数分裂(如样本12-1与11-34)。
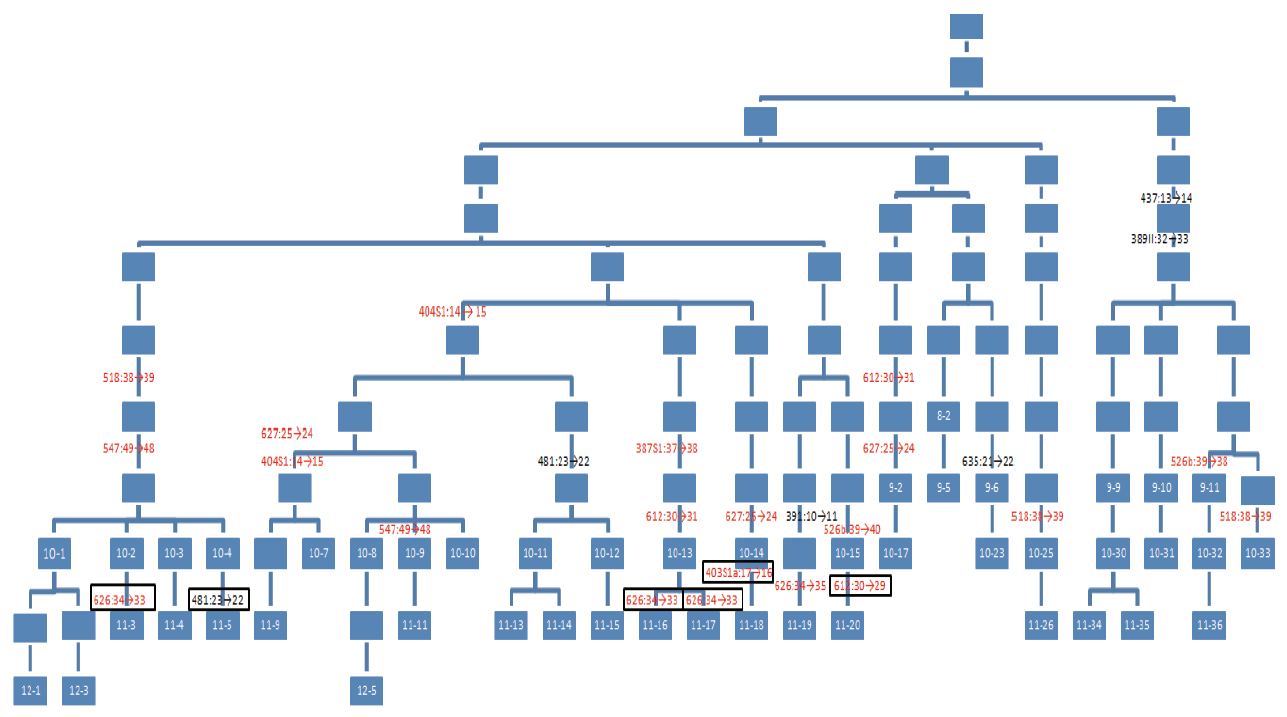 | 图1 家系图谱以及突变位置定位(有编号的蓝色方框代表实验样本, 其他为逝者或未能采集到的样本; 表示准确的突变位置, 其他均为推测的突变位置; 红色:RM Y-STRs; 黑色:其他Y-STRs)Fig.1 Lineage map and mutations localized (Blue bars with serial numbers: tested samples; the others: deceased or unavailable representatives. indicting the precise site of mutation; the others: the deduced site of mutation. Red: RM Y-STRs, Black: other Y-STRs) |
ABI9700型PCR扩增仪; ABI3500xL型遗传分析仪; MagAttractM48 DNA Manual Kit(Qiagen); Yfiler® Plus试剂盒; 2× PCR Master Mix (Promega); GeneScan600 LIZ Size Standard; Hi-DiFormamide (Applied Biosystems)。
使用M48试剂盒提取样本DNA。使用Yfiler® Plus试剂盒和7个RM Y-STRs复合扩增体系对DNA进行扩增。试剂盒的具体操作均参照产品说明书。7个RM Y-STRs复合扩增体系(10 uL)包括:5 μ L 2× PCR Master Mix (Promega), 4 μ L Primer Mix (引物序列参照Ballantyne等[2], 终浓度为0.4 μ M), DNA模板1μ L。PCR程序如下:95 ℃、2 min; 28个循环(94 ℃、30 s; 59 ℃、40 s; 72 ℃、40 s); 72 ℃、20 min。PCR产物在ABI3500xL遗传分析仪上进行检测。
7个RM Y-STRs复合扩增体系中等位基因的命名:对样本10-1每个基因座的扩增产物进行测序, 根据核心序列的重复次数对其等位基因分型进行命名; 对于其他样本, 减少一个重复序列长度的等位基因的命名在10-1的基础上减1, 增加一个重复序列长度的等位基因的命名在10-1的基础上加1。根据样本之间的亲缘关系和Y-STR分型数据, 在家系图中定位突变发生的位置。
突变位置的定位遵照以下原则:1)若父子之间某Y-STR基因座分型不一致, 则突变位置可以直接定位在父子之间的传代过程; 2)若家系中某一分支的个体在某基因座上与其他分支人员不一致, 考虑到突变的稀有性, 认为在这个分支与其他分支分离的部位发生了突变。
比如, 样本10-9和11-11 DYS547的分型为48, 而10-8、10-10、12-5DYS547的分型均为49, 最合理的解释是样本10-8、10-9、10-10的父亲的DYS547的分型应为49, 只是在传递给10-9的过程中突变为48; 再比如10-11、10-12、11-13、11-14、11-15的DYS481分型为22, 而与这个分支临近的其他样本均为23, 最合理的解释是10-11和10-12所在的这个分支发生了突变, 可能是在10-11与10-12的祖父传递给10-11与10-12的父亲时发生了突变, 也可能是其曾祖父传递给其祖父时发生了突变, 只有这样才能通过一次突变导致这个分支的所有成员的DYS481分型与其他分支不一样。
根据突变定位的结果统计Y-STR基因座的突变次数; 由于样本较少, 涉及到的减数分裂次数不足(106次), 未分别计算每个Y-STR基因座的突变率。统计家系中Yfiler、YfilerPlus和RM Y-STR 3种Y-STR单倍型的个数并比较两两个体之间的最大差异基因座个数和单倍型差异率(Rate Referred to Haplotype Differences, RRHD)[6]。RRHD指家系中具有特定亲缘距离(减数分裂次数)的两名男性个体之间单倍型不一致(≥ 1个基因座的分型不同)的概率, 表示为单倍型不一致的样本对数/总样本对数。
在7个RM Y-STR复合扩增体系中, DYS399S1(三拷贝Y-STR基因座)等位基因峰图扩增不平衡现象较为明显, 且部分样本检测出4至5个等位基因, 分型判断难度较大, 因此, 数据分析剔除了DYS399S1基因座。在剩余的31个Y-STR基因座中, 14个Y-STR基因座检出1个以上的等位基因, 推测共发生了27次突变, 其中DYS626突变4次, DYS518、DYS612和DYS627各突变3次, DYF404S1、DYS481、DYS526b和DYS547各突变2次, DYF403S1a、DYF387S1、DYS389II、DYS437、DYS391和DYS635各突变1次。其余17个Y-STR基因座未发现新等位基因, 认为没有突变发生(未考虑传代过程中可能发生的回复突变)。突变发生的位置(或可能的位置)见图1。在27次突变中, 9个RM Y-STR (DYS626、DYS518、DYS612、DYS627、DYF404S1、DYS526b、DYS547、 DYF403S1a、DYF387S1)突变21次, 占77.8%; 另外5个Y-STR (DYS481、DYS389II, DYS437, DYS391, DYS635)突变6次, 仅占22.2 %。减少重复单位的突变为13次, 占48.1 %, 增加重复单位的突变为14次, 占51.9 %。27次突变均为1步突变。全部31个基因座的平均突变率为7.08‰ (95% CI, 4.7‰ ~10.3‰ ), 12个RM Y-STR基因座的平均突变率为12.4‰ (95% CI, 7.7‰ ~18.9‰ ), 其余19个基因座的平均突变率为2.83‰ (95% CI, 1.0‰ ~6.2‰ )。
在47名男性家族成员中, 发现4种Yfiler单倍型, 12种YfilerPlus单倍型, 16种RM Y-STR单倍型, 对于全部31个Y-STR基因座, 发现21种单倍型(表1)。
| 表1 47名男性个体的Y-STR单倍型 Table 1 Y-STR haplotypes of 47 male individuals |
对于Yfiler单倍型, 两两个体之间最大差异基因座数为3个(如样本9-9与11-19); 对于YfilerPlus单倍型, 两两个体之间最大差异基因座数为4个(如样本9-2与11-5); 对于RM Y-STR单倍型, 两两个体之间最大差异基因座数为5个(如样本10-13与11-3); 对于31个Y-STR组成的单倍型, 两两个体之间最大差异基因座数为6个(如样本10-32与11-3)。47名个体两两之间的亲缘距离介于1次减数分裂到19次减数分裂, 样本对数为13至134对, 共计1081个样本对。两两个体之间的RRHD以及最大差异基因座数见表2。当两名个体相隔一次减数分裂(父子)时, 计算RRHD, Yfiler单倍型为0, YfilerPlus单倍型是4.76%, RM Y-STR单倍型则为23.8%, 全部31个Y-STR的RRHD为28.57%; 当两名个体相隔10次减数分裂时, 对于RRHD, Yfiler单倍型为7.35%, YfilerPlus单倍型为91.18%; RM Y-STR单倍型和全部31个Y-STR组合值为100 %。
| 表2 三个Y-STR分型体系的单倍型比较 Table 2 Comparison of haplotypes obtained from three different Y-STR kits |
Y-STR单倍型中Y-STR基因座数量越多, 在同一家族中单倍型产生差异的概率就越大。在本研究中, 同一家族中47名男性个体发现了4种Yfiler单倍型, 12种YfilerPlus单倍型, 16种RM Y-STR基因座, 另外, Y-STR的突变率对单倍型的变异程度影响较大, 比如12个RM Y-STR组成的单倍型的差异程度超过了25个Y-STR组成的YfilerPlus单倍型。这提示我们, 当分型体系中Y-STR个数较多或者含有RM Y-STR基因座时, 排除家系一定要慎重。当前, 广泛应用的Y-STR复合扩增体系包含Y-STR基因座越来越多, 并且引入了部分的RM Y-STR基因座, 这在增加单倍型多样性的同时也增加了误排父系亲缘关系的风险。对于16个YfilerY-STR基因座, 一般认为, 3个及以上基因座的不一致可以排除父子关系[8]; Liu 等[9]提出, 对于Yfiler单倍型, 不超过2个基因座不一致(累计突变步数不大于2步)提示两名个体来源于同一家系的可能性非常大, 对于YfilerPlus单倍型, 这一标准为不超过4个基因座不一致(累计突变步数不大于5步)。我们认为, 上述标准仍有适当放宽的必要, 特别是在同一家系中的两人亲缘距离较远的情况下, 比如本研究中就发现了两名个体17个Yfiler Y-STR基因座中有3个基因座不一致的情况(表2)。另外, 在基因座数量进一步增加特别是分型体系中包含RM Y-STR的情况下, 排除标准需要根据具体情况进行分析, 综合考虑突变的性质(突变基因座的个数、突变率、突变步数等), 可以预见, 如果个体之间亲缘距离更远, 或者Y-STR基因座的数量继续增加, 分型不一致的基因座的数量仍将继续增加。
家系内男性个体之间的RRHD随着减数分裂次数的增多和Y-STR基因座数量的增加而逐渐增大, 这也提示我们, 增加Y-STR基因座特别是RM Y-STR基因座的数量可以在家系内部进一步细化区分亲缘关系, 甚至为利用Y-STR单倍型进行个体识别提供了可能; 本研究中, 当两名个体相隔10次减数分裂时, YfilerPlus单倍型的差异率为91.18%, RM Y-STR单倍型差异率则达到了100%, 甚至有23.8%的父子对个体的RM Y-STR单倍型存在差异, 这与Ballatyne 等人的研究结果一致[10]。同时, Y-STR数量的增加和RM Y-STR的应用使得父系亲缘关系判定更为复杂, 在实践中需要权衡利弊, 根据需要选用不同的Y-STR分型体系。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|